Cino
EA, Karttunen M, Choy WY (2012) Effects of molecular crowding on the dynamics
of intrinsically disordered proteins. PLoS One 7:e49876. link to
manuscript
Inside cells the concentration of macromolecules can reach up to 400 g/L, creating a crowded environment (Fig 1). The space occupied by cellular molecules (proteins, nucleic acids, etc) reduces the amount of water available, causing molecules to behave differently than they would in more dilute environments. Most studies of proteins and other macromolecules are conducted in vitro with purified and relatively dilute samples. To accurately characterize macromolecules and the biochemical processes they are involved in, it is important to examine them in vivo, or under conditions that mimic the crowded cellular environment.

In the crowded cellular environment, proteins are expected to behave differently than in vitro. The stability and the folding rate of a well-folded protein can be altered by the excluded volume effect produced by a high density of macromolecules. However, crowding effects on intrinsically disordered proteins (IDPs) are less explored. These proteins can be extremely dynamic and potentially sample a wide ensemble of conformations (Fig 2). The dynamic properties of IDPs are intimately related to the timescale of conformational exchange within the ensemble, which govern
target recognition and how these proteins function.
In this manuscript, we focused on determining
how molecular crowding affects the dynamics of IDPs using NMR spin-relaxation experiments.
Measurements were taken for three disordered proteins, and the well-folded
protein, ubiquitin, for comparison, in the absence and presence of crowding
agents. Our data illustrates that IDPs remain at least partially disordered
despite the presence of high concentration of other macromolecules (Fig 3).
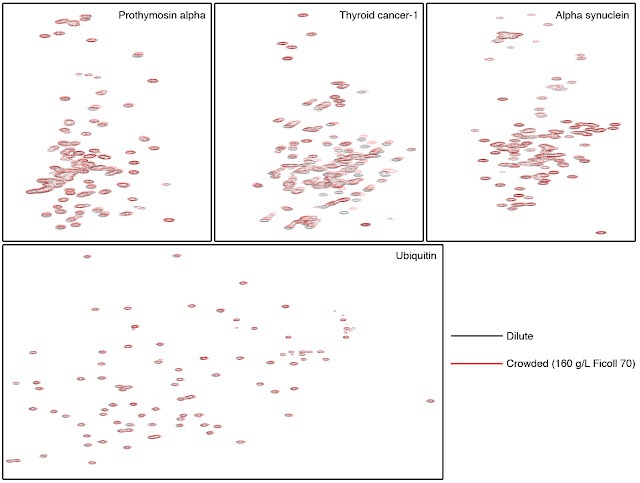
Fig 3. 1H-15N Heteronuclear Single Quantum
Coherence (HSQC) spectra in the absence (black) and presence (red) of 160 g/L
crowding agent Ficoll 70. The IDPs Prothymosin alpha, Thyroid cancer-1 and
alpha synuclein as well as the well-folded protein, ubiquitin, were examined.
Similar black and red spectra indicate that the protein structures are similar
in dilute and crowded environments.
Despite this, specific regions of Thyroid-cancer-1 and Prothymosin alpha, which encompass protein-protein interaction sites exhibited differential dynamics in the absence and presence of high concentration of crowding agents (Fig 4). This suggests that the crowded environment may have differential effects on the conformational propensity of distinct regions of an IDP, which may lead to selective stabilization of certain target-binding motifs.
Despite this, specific regions of Thyroid-cancer-1 and Prothymosin alpha, which encompass protein-protein interaction sites exhibited differential dynamics in the absence and presence of high concentration of crowding agents (Fig 4). This suggests that the crowded environment may have differential effects on the conformational propensity of distinct regions of an IDP, which may lead to selective stabilization of certain target-binding motifs.
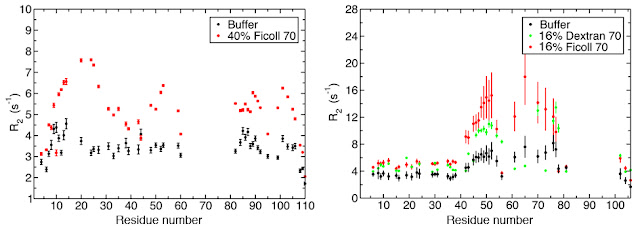
Fig 4. Backbone N-H bond transverse relaxation rates for prothymosin
alpha and thyroid cancer-1 in the absence (black) and presence (red and green)
of crowding agents. Distinct regions of the proteins show differential changes
in dynamics in response to crowding.
Using an MD simulation of prothymosin alpha in the absence of crowding agents, we have proposed a model to correlate the observed changes in relaxation rates to the alteration in protein motions under crowding conditions (see the manuscript for details). Overall, the results show that the segmental motions of IDPs on the nanosecond timescale are retained under crowded conditions and that IDPs function as dynamic structural ensembles in cellular environments.
Using an MD simulation of prothymosin alpha in the absence of crowding agents, we have proposed a model to correlate the observed changes in relaxation rates to the alteration in protein motions under crowding conditions (see the manuscript for details). Overall, the results show that the segmental motions of IDPs on the nanosecond timescale are retained under crowded conditions and that IDPs function as dynamic structural ensembles in cellular environments.
Our related work references
1.
Cino EA, Karttunen M, Choy WY (2012) Effects of molecular crowding on the
dynamics of intrinsically disordered proteins. PLoS One 7:e49876. link to
manuscript
2.
Cino
EA, Choy WY, Karttunen M (2012) Comparison of Secondary Structure Formation
Using 10 Different Force Fields in Microsecond Molecular Dynamics Simulations. J
Chem Theory Comput 8:2725-2740. link to
manuscript
3. Cino EA, Wong-Ekkabut J, Karttunen M, Choy
WY (2011) Microsecond molecular dynamics simulations of intrinsically
disordered proteins involved in the oxidative stress response. PLoS One 6:e27371.
link to
manuscript
4.
Cino E, Fan J, Yang D, Choy WY (2012) (1)H, (15)N and (13)C backbone resonance
assignments of the Kelch domain of mouse Keap1. Biomol NMR Assign. In press. link to manuscript
5. Khan H, Cino, EA, Brickenden A, Fan J, Yang D, Choy WY (2013)
Fuzzy Complex Formation between the Intrinsically Disordered Prothymosin α and the Kelch Domain of
Keap1 Involved in the Oxidative Stress Response. J Mol Biol 6:1011-1027. link to manuscript